"Avian Flight"より抜粋
(始祖鳥はバシリスク型の水上走行者であったか?)
John J.Videler
5.4 「イエス=キリスト恐竜」理論
中米のトカゲBasiliscus属は,縮小した恐竜に似ている.彼らの俗名の1つは,水面を走って渡る能力に由来する「イエス=キリスト トカゲ(Jesus-Christ lizards)」である.(Deventer 1983) (Basiliscus属の)4種は全て2足の疾走者である.小さな個体は捕食者から逃れるため,そして新たな摂食エリアを利用するために,水溜りの向こう側に走り抜ける(図5.3(a)).成体の体重は200[g]から600[g]の間で様々である.雄は最大で体長1[m]に達し,雌は体長0.6[m],体重300[g]にまで成長する.彼らの体長の4分の3は尾で占められる.このトカゲは,長くてやや平らな足指を伴う長い後肢を持っている.脚部が水面を叩いたとき,足指は回転する.回転の後に,爪先に沿った側面のフリンジが,面積を増やす(Laerm 1974).最大走行速度は2.3[m/s]が報告されている(Rand and Marx 1967).
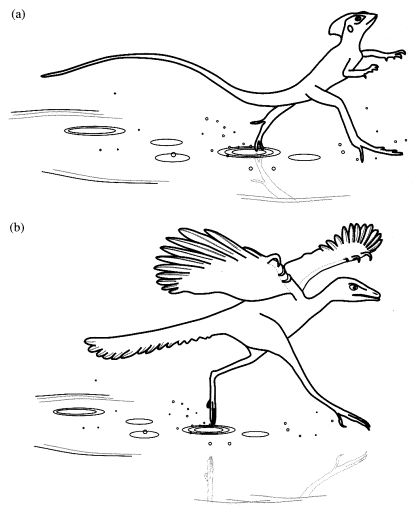
Fig.5.3 水面を走る「イエス=キリスト トカゲ」(a)そして始祖鳥(Archaeopteryx)(b).トカゲの描写は,表紙絵として使われたスティーヴン・ダルトンによる写真に基づいている(Alexander 1992).始祖鳥(Archaeopteryx)の相対的な寸法はベルリン標本に基づく(Videler, 2000).
水上を走る間の体重の支持は,表面張力とは何ら関係はなく,水面を叩くことによって動的に達成される.各ステップの間に,3つの段階が顕著となる.「打撃段階」では,扁平な足が水面を打つ.その後,「ストローク段階」では,水中に空気による空洞(air cavity)を作り出すために押し下げる.「引込段階」では,空気の空洞が崩壊する前に,足はすばやく引っ込められる.この種の歩行運動の有効性を決定する上で,脚部のサイズと下腿の長さは重要な要因である.
水面を走る能力はサイズに依存する──200[g]のトカゲはかろうじてその体重を支えることができる.大きさと速度の間には相関が無いが,大きな成体は幼若体ほどには遠くまで走ることができない.歩行周期は5~10[Hz]の様々である──常に一方の脚が水中にあるため,歩みの区切りは0.1~0.05[s]で変化する.重い成体の雄は水面を打つときに深く沈みすぎ,空気の空洞(air cavity)が崩壊する前に抜け出すには引き込み速度が十分ではない.結論として水面を走る能力は,体重,歩行周期,そして(水面を)叩く脚部の速度に依存する.
私の仮説によれば,始祖鳥(Archaeopteryx) の先祖は,水面を走るトリックを利用して肉食動物から逃れ,1億5000万年前の中欧のサンゴ礁で島の間を移動する,「イエス=キリスト恐竜」であった(Fig. 5.3(b)).これらの先祖はすでに羽毛を持ち,そしておそらく水の上に浮かぶことができた.最初の段階では,彼らは前部の手足を水上で走っている間に平衡状態を保持するために使った.
より重い体を補い,水上走行距離を伸ばすために,次第に腕は揚力を生成する翼に進化した.はじめは推力と体重の支持は両者ともに水面を叩く脚部で提供され,後に主翼が若干の体重支持を受け持つようになった.すべての小さい段階が揚力の増加を指向するこのシナリオは,より優れた逃走能力とより広い摂食領域を提供し,動物の適応度に対して肯定的な効果があった.
自然淘汰は翼の揚力を生成する性質を最適化するために働いていた.始祖鳥(Archaeopteryx)の翼(我々は化石証拠からそれを知っている) は,脚で作り出せる程度の低速において,羽ばたかずに揚力を生み出すことに適応していたが,まだ推力を生み出すために使うことはできなかった.
ロンドン,ベルリン,ミュンヘン,そしてアイヒシュテット標本は,水面を叩くために必要とされる運動を可能にするのに十分な,90°以上に脚部を曲げることができることを示す.始祖鳥(Archaeopteryx) は取り付けられたフリンジを持っていたかもしれないが,その軟部組織の化石化した痕跡はこれまでのところ識別されていない.後肢の解剖学的な態様は,バシリスクが行う脚の折りたたみ動作を始祖鳥(Archaeopteryx) が行えないという仮説について,明白な理由を示さない.水面上を走ることは,足関節の強さと大きな動きの両方を必要とする.始祖鳥(Archaeopteryx) は,バシリスクの持つこれらの特徴を共有して持っている.
始祖鳥(Archaeopteryx)の翼はRietschelによって詳細に研究された.彼の右翼の復元をFig. 5.4に示す.(Rietschel, 1985)翼はその大きさの鳥類としては例外的に大きく,このような重々しい造りの翼をはばたくには多くの力を必要とするであろう.手の部分は主翼全体の長さのおよそ40%を占める.
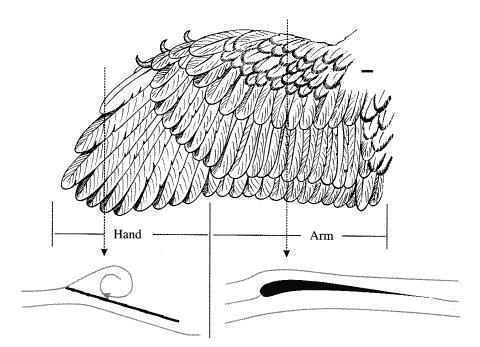
Fig.5.4 Rietschelによる,腹側から見た始祖鳥(Archaeopteryx)の右翼の復元.(著者の好意による許可(に伴う掲載);スケールバーは1[cm]) 主翼の手と腕の部位を通しての断面図は,揚力の発生を予期させられる流れのパターンを示す.(Videler 2000)
腕の部位は十分に反っている.腕を通る断面図は丸められた前縁(leading edge),大きい翼弦(※原文ではcordだが,文脈的にchordの事であろう)と鋭い後縁を見せたであろう.明確に生成された腕の翼は高迎角(high angles of attack)においてさえ,定常的に揚力を発生する.手の部位は,羽毛の前縁を形づくる幅の狭い羽根を伴う12の初列風切羽から成り立つ.手の翼の前縁は鋭く,後ろ向きには楔状であり,初列風切羽の羽軸は少し後方に湾曲している.事実上,手の翼は鋭い前縁を持つswept-backのデルタ翼である.
動的な失速を遅延させる低速かつ高迎角は,特に低速において60~70°程度の高迎角に対処するために適応した形態(configuration)の主翼(第1章および4章を参照)の上に,前縁渦を作る.機構上の欠落により,始祖鳥(Archaeopteryx) はその翼をはばたくことができなかったが,おそらく様々な速度で必要とされる揚力の総量を制御するため,扇のようにその翼を展開し,閉じることができた.この概念は3本の指の鉤爪に明確な役割を提供する.
近位の翼部の付着流(attached flow)が,sharp-edged でswept-backな遠位の翼部の上の分離された前縁渦(leading edge vortices)と同じくらい良く揚力を生成するように航空機は設計されており,そのために,しばしば2種類の流れを分離する装置が要求される.境界層板(wing fences)と鋸歯翼(saw-toothed leading edges) (Fig. 5.5a)は,境界層をきれいにし,次に来る一連の流れを分離する渦を作るために設置される.腕と翼端部の間に位置する指の爪は,航空機において前縁渦(leading edge vortices)の態様を制御するのと同様に役立つ可能性のある前縁突出部(leading edge protuberances)である.
(Ashill et al. 1995' Barnard and Philpott 1997; Lowson and Riley 1995; Videler 2000).
コウモリの前縁鉤爪(図5.5b),数種の翼竜(図5.5c),そして鳥類の小翼(alula)はおそらく同じ機能を持っていた.
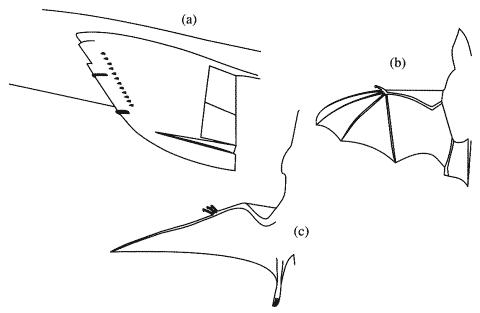
Fig.5.5 主翼の前縁機構. (a)ブリティッシュ・エアロスペース・ハリアー戦闘機の境界層板(wing fences)と鋸歯翼 (b) コウモリ翼の第一指 (c) プテラノドン(pteranodon) の自由に動く3本の指.
始祖鳥(Archaeopteryx)の鉤爪はおそらく動的に最適な方法でその仕事を行なった.3本の鉤爪のそれぞれは,急速な遷移が必要なときでさえも,手羽先の翼(面積)の総量に従って使うことができた.使用中でない鉤爪は羽毛の間に引込んで置かれることがあり得た.
この機能は,鉤爪はなぜ磨耗を示さないのか,そしてなぜbi-stable関節(bi-stable joint)が最終指と残りの指の結合を形成するのかについて,説明するだろう.
この特徴のために,鉤爪は2種の既定位置を得た──伸展時と,格納時である.
尾部は後ろ向きに真直ぐな状態に保たれ,水上走行の制御と安定のために,さまざまな量の揚力と抗力を生成することができた.わずかな走行移動に使われた低速において,それは制御に致命的な影響は与えていないだろうが,それは推進時に足で水面を連打する上で,動物を良好なバランスに保つことの助けになったであろう.
始祖鳥化石がほとんど宗教的と言うべき地位(Witmer, 2002)を持っているため,一般に,始祖鳥(Archaeopteryx) に関する新しい学説は極端な懸念を抱いて扱われる.学術誌「Archaeopteryx」における論文(Ma et al. 2002)は,同じ学術誌で2000年に発表した私の説に疑義を提起し,それが誤りであることを証明しようとした.次項にそれに対する反論を示す.
5.5 始祖鳥(Archaeopteryx)はいかにして水上で走ることができたか?
この疑問に答えるためには,力のバランス関係についての量的な評価が要求される.Glasheen とMcMahonによる量的な生体機構的なモデル(1996a,b)は,水上走行におけるバシリスクの振舞いを説明する.これらのモデルはサイズに依存した重要なパラメータを示すアロメトリー方程式(allometric equations)を提供する.モデルは箱5.1に説明される.
この手法は,脚で水面を叩くことで始祖鳥(Archaeopteryx)が作り出すことができたはずの推進力を予測する機会を提供する.各ステップで体重が要求する最小の衝撃力は,打撃段階およびストローク段階(slap and stroke phases)で作られる縦方向の衝撃力の合計よりも小さくなければならない.主翼と尾からの滑空による総揚力は,始祖鳥(Archaeopteryx) の体の重さを減少させた.
揚力とそれに伴う誘導抵抗(induced drag)は,第4章の質量流束モデルの修正版を使って見積った.疾走する動物の主翼と尾部によって下方に逸らされた空気の質量は考慮に入れられる.そのモデルを記述している方程式を箱5.2に示す.こころもち斜めのストロークの間に脚部によって産み出された推進力は,それぞれのステップの間において,抗力の見積りを超えるべきである.力のバランスの結果は,始祖鳥(Archaeopteryx) に水上を走ることが果たして物理的に可能であったか,という疑問の答えとなるだろう.
(表5.1はそれぞれのモデルにおいて使われた物理量を示す.)


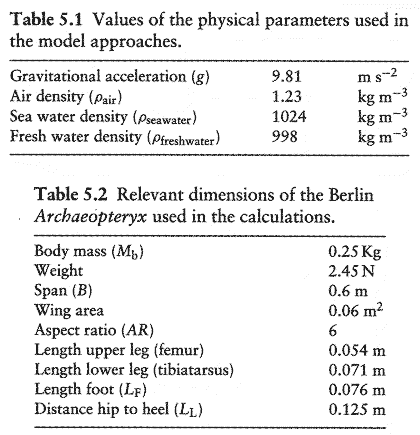
表5.1 本モデルアプローチに使用した物理パラメータの値.
表5.2 計算に使用された,始祖鳥ベルリン標本の関連寸法.
5.5.1 脚部と水の間の相互作用の計算
下腿の骨格の要素の長さの測定はベルリン化石(図5.1)から直接行われた.主板(それは動物の左側であったはずだが,どのように化石化したかについてはまだ論争がある)の上に現われる右脚はほとんど完全に維持されており,その全ての長さについて同一面上に位置しているため,補正なしでそれを測定することができる.Yalden (1984)は,始祖鳥(Archaeopteryx) の体重範囲の学問的な推測を立てた.0.25[kg]という値は,ベルリン標本について適正で正確な見積りであると思われる.始祖鳥(Archaeopteryx) ベルリン標本の諸元を表5.2に示す.
始祖鳥(Archaeopteryx) の下腿と脚部のサイズは,Glasheen とMcMahonのアロメトリー方程式(allometric equations)を使って,同サイズのバシリスクと比較される.(箱5.1)バシリスクの下腿と脚部の長さは,体重に対するほぼ100倍の範囲に近く位置づけられる.始祖鳥(Archaeopteryx) の足先(foot)はバシリスクより短いが,全体の脚長はより長い.
バシリスク・モデルはそれぞれの行程を前に言及した3つの段階に分ける:(1) 打撃段階,(2)ストローク段階 (3)延長段階(protraction)である.最低限必要な縦方向の([Ns]での)衝撃は,体重とステップ持続時間の積である.常に片脚が水中にあり,それぞれの歩みが2ステップの期間を要する.我々は,始祖鳥(Archaeopteryx) について「片脚が水中に無い滑空段階」を無いものとする条件を受け入れる.打撃は垂直方向に向けられると想定する.
最大打撃力は,水の密度,打撃速度,そしてGlasheenとMcMahon.によって試されたバシリスクの足の物理モデルと同じように生成される円弧の半径上の第3の力に比例する.バシリスクの打撃速度は極めて変動的であり,体の大きさの関数ではなかった.
GlasheenとMcMahonは上限として3.75[m/s]を使った.これは行程の間に測定した下方速度の二乗平均平方根値(root mean square value)の1.5倍である.これらの値が始祖鳥(Archaeopteryx)に使えないと想定する明白な理由はない. 最大行程衝撃力は,下降する脚の平均抗力と,抗力が作用している時間の産物である.
このモデルは,垂直下方に近い一定の速度でのストロークの深さは,下腿の長さと等しいと想定する.脚は全ての行程の間,進行の方向に対して直角の状態に保たれる.ストロークは,わずかな小角β分だけ後方にある.最大ストローク衝撃力の垂直および水平成分は,余弦βと正弦βのそれぞれの乗算によって導かれる.
垂直方向の最大ストローク衝撃力と最大打撃力の合計は,水上走行を可能とする衝撃力の最小要求値よりも大きくなければならない.ストローク衝撃力の水平成分は,走行速度で抗力に打ち勝つために必要な衝撃力よりも大きくなければならない.
始祖鳥(Archaeopteryx) の潜在的な揚力生成能力は,表5.3において紹介された結果では考慮に入れられていない.Glasheen とMcMahonは,5Hzと10Hz の間の歩み頻度がバシリスク(Basilisk lizards)に使われ,体重に対する明確な依存はないことを発見した.
従ってこれらはここで同様に使われる.真水より高い密度を持つ海水を叩いていたという事実を踏まえるなら,始祖鳥(Archaeopteryx) の最大衝撃力はわずかに高い.
始祖鳥(Archaeopteryx) のより長い足は,おおむね(バシリスクよりも)高い最大行程衝撃力を発生する.10[Hz]の歩み頻度(0.05[s]のステップ持続時間)における全体での最大衝撃力は,両方の動物において必要とする最小衝撃を超える. 始祖鳥(Archaeopteryx) における衝撃力の余剰分は,脚長の相違の結果として,同じ体重のバシリスクのそれより大きい.
わずか5[Hz]の歩み頻度の衝撃余剰では,始祖鳥(Archaeopteryx) とバシリスクの両者ともに水面を走れないことを意味する.
0.25[kg]のバシリスクが(水面走行の)トリックを行なうためには,9.2[Hz]の最小歩み頻度を必要とすると,このモデルは予測する.始祖鳥(Archaeopteryx) は所要な衝撃を減らすための揚力を生成する可能性を持っている.
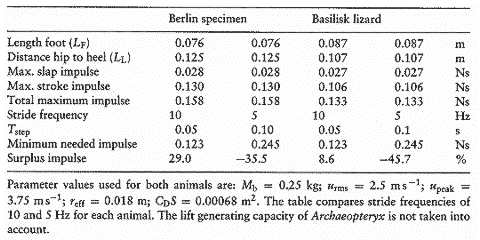
表5.3 始祖鳥(Archaeopteryx)とバシリスクの水上走行能力の比較.
5.5.2 走る始祖鳥(Archaeopteryx) によって生成される空気力(aerodynamic force)の見積もり
始祖鳥(Archaeopteryx)の大きな主翼と羽がついた尾部は,水上を走る間に流れてくる空気を下方にそらすことにより,揚力を生み出した. 空気と主翼および長い横向きの羽がついた尾部の間の干渉は,どちらかと言うと複雑であったに違いない.このモデルは,影響を受けた空気を同一の流れの円柱形の空気によって表せると想定することによって,その複雑さを単純化する.
図5.6は動物が一定な速度においてどのように走っているか,広げられた翼と展開された尾翼の存在のために,円筒形の空気がいかにして下方にそらされるかを示す.
円柱の直径はおおむね翼幅と等しいと推定される.
第4章(Box 4.1)において,理論を記述したマス・フラックス法(mass flux method)は,一定の速度での走行において始祖鳥の翼と尾が作り出すことができる揚力の総計の推測することに適用できる.
アレクサンダー(Alexander, 1976)によって見積もられた,始祖鳥(Archaeopteryx)サイズの恐竜の最大2足走行速度2[m/s]が,計算に使用される.表 5.2は計算において使われた方程式を示す.単位時間当たりに関係する空気の体積は,円柱断面に走行速度を乗算した値と等しい.大気密度による乗算は単位時間当たりの空気の質量に用いられ,それは空中で力が用いられていることを見いだすための下方への速度で乗算される.疾走する動物は,揚力を発生するために,その主翼と尾部によって吹降し角(downwash angle) 8°以上で空気を押し下げることを推進する.揚力は空気を押し下げることを促進することによって動物に働く反作用である.
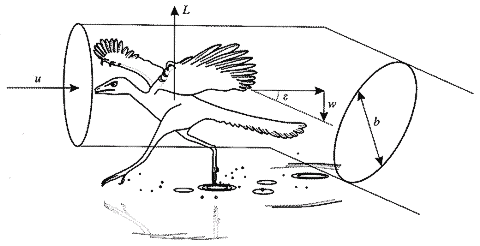
Fig.5.6 水上を速度vで走る始祖鳥(Archaeopteryx)についてのアーティストのイメージ.
動物に対して,気流は速度vを持っている.
滑空位置にある翼,体,尾部の統合された動きにより,気流は8°以上下方へそらされる.
翼幅と等しい直径bの空気の円柱領域が影響を与えられる.
空気は下方へ加速され,速度wに到達する.航空力学的反作用力Lは上方に向けられる.詳細については本文を参照のこと.
この動きは動物の上に走行している反対方向の抵抗力を誘発するであろう.誘導抵抗(induced drag)と走行速度の乗算したものが誘導力(induced power)である.誘導力は下方の空気に与えた一定時間の運動エネルギーに等しい.誘導抵抗(induced drag)は誘導力(induced power)を走行速度で除算することで導くことができる.
動物に働く抗力は,揚力の発生と関連する誘導抵抗(induced drag),翼と尾の形状抵抗(profile drag),そして頭,体,脚に働く有害抵抗(parasite drag)から成り立つ.形状抵抗(profile drag)は,気流が下方に逸らされない位置を維持するとき,翼と尾に働く抵抗である.有害抵抗(parasite drag)は,大気中を移動する間に,体のそれ以外の部分に働く圧力および摩擦に結び付けられる.
形状抵抗(profile drag)と有害抵抗(parasite drag)は算定することが難しい.これらは,揚力を生成せずに走行している間は前面投影面積(frontal area)に比例し,より効果的に気流を阻止する面積の割合である抗力係数に比例する.
このモデルは,動物の背後に生じる気流の方向と速度についての妥当な推測を用いることで,動物と気流の干渉を統合する.揚力作用の見積もりは,翼,体,尾部の統合化された効果を含み,腕の翼と手羽先が異なった方法で揚力と抵抗を生成するという可能性は考慮しない.地面効果によって高められた揚力を利用する上で(第4章を参照),主翼は十分に水面に近い.地面効果の大きさは,揚力に比例し,翼のアスペクト比(翼幅の二乗を翼面積で除算したもの)に反比例する(Anderson および Eberhardt, 2001) (図5.7).
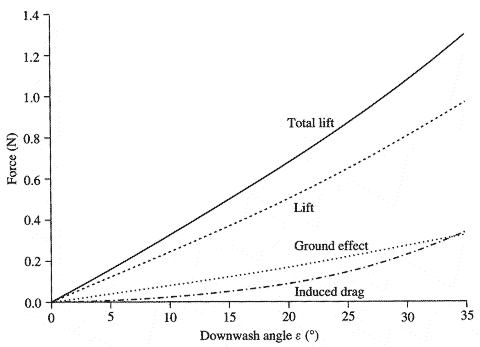
Fig.5.7 2[m/s]で水上を走る始祖鳥(Archaeopteryx)が受ける揚力と抗力のダウンウォッシュ効果.力は,SunadaおよびEllington(2000)による質量流束モデル法(mass flux model approach)に基づいて計算された.詳細については本文を参照のこと.
0~35度の間における8種類の吹降し角(downwash angles)についての空気力(aerodynamic force) の計算結果を,図5.7に示す.円柱形の仮想的な空気の質量を逸らすことにより生成される総揚力は,吹降し角(downwash angle)に相関して増加する地面効果(ground effect)によって増加する.2[m/s]で疾走する始祖鳥(Archaeopteryx) に働く誘導抵抗(induced drag)は同様に増加するが,それは動物を持ち上げる揚力の値と比較して小さいままである.図5.8は,吹降し角(downwash angles)の範囲における揚力がどのように正味の衝撃余剰を決定するか,そしてそれにより3つの歩み頻度で水面を走る能力を示す.すでに我々は,0.05[s]の歩幅周期ならば,始祖鳥(Archaeopteryx) が揚力の手助けなしでも水上を走行可能であることを確認している.0.1[s]の歩幅周期では,0.16[Ns]以下の衝撃分の力として,主翼・尾翼からの揚力を要求する.吹降し角が26度を超える場合の我々のモデルによれば,その必要分は生成されるであろう.0.07[s]の場合における平均ステップ周期の結果は,同じく図5.8に提示され,このケースでは約11度以上の吹降し角(downwash angles)が必要であることを示す.
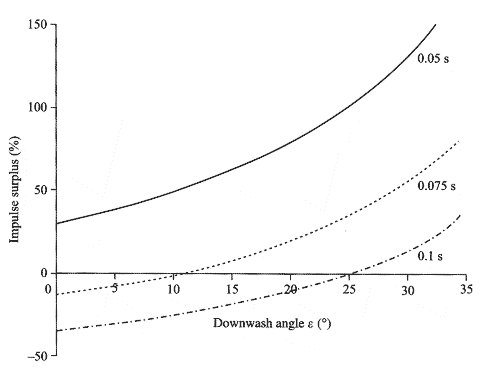
Fig.5.8 2[m/s]で水面を渡って走行している間,バシリスク(Basilisk lizard)の技法を使うとともに,安定した翼と尾で気流を下方に誘導し,揚力を作り出すことで,0.25[kg]の始祖鳥(Archaeopteryx)における衝撃の余剰分は生成されるであろう.モデルは3つの段階の持続時間についての予測を示す.
形状抵抗(profile drag)と有害抵抗(parasite drag)の合計は,0.009[m^2]の前面投影面積の見積りと0.25の(空力)係数を使った場合には0.006[N]のオーダーであると推定され,これは無視できるほどに十分に小さいと考えられる.
ε= 26"における0.165 [N]の誘導抵抗を克服するため,それぞれのステップの間に作り出されなければならない衝撃の前方成分(forward directed impulse)は,0.017 [Ns]である.
この値はわずか7.5度の戻り行程角度βにおいて達している──衝撃の縦方向成分(vertical component of the stroke impulse)の0.130 [Ns]のうち0.129[Ns]を残しており,余剰分の割合はこの程度の小さな戻り行程角度ではほとんど影響を受けないことを示す.
10[Hz]程度の走行周期を持つか,揚力を生成することができたならば,0.25[kg]の始祖鳥(Archaeopteryx)は水面を走ることが可能であったことを,分析結果は示している.
*
原文:
John J.Videler. 2005. "Avian Flight", pp. 102-114.
OXFORD ORNITHOLOGY SERIES,
OXFORD UNIVERSITY PRESS,
ISBN 0-19-929992-7
|